
INTRODUCTION
About 8 years ago I was studying genetics in detail to see how the hand of God worked at the DNA and cellular level. As part of that research I read several amazing scientific books, Genetic Entropy and The Cell’s Design. Their insights were so insightful that I summarized their contents for a wider audience. You can find the links for these summaries under the Genetic Entropy tab and the Science tab on the front page of my website.
However, in the 8 years since that research an avalanche of new knowledge has emerged. This essay is a summary of my latest reading and research into the human genome from two recent books, Replacing Darwin and Traced, both by Dr Nathaniel Jeanson, which have brought me up to date in this fast-moving field. They have shone an even more powerful light on our true human origins, so I have penned for you a simplified layman’s glimpse of the insights into our recent creation and the accuracy of Genesis that I gleaned from these two books. I will only be using evidence from DNA and nothing else.
Over 160 years ago Charles Darwin’s radical book, The Origin of Species, transformed our intellectual and cultural world forever, even though most people alive today have never read it. The book very astutely drew on data from ecology, palaeontology, geology, biogeography, anatomy, physiology, and embryology to argue the case for the unguided evolution of life over eons of time. Within two decades of its 1859 publication the academic world was on Darwin’s side as it completely destroyed the then creationist position at the time of the fixity of each of today’s species at the original creation.
However, Darwin’s seminal work had a massive blind spot. It never actually dealt with the scientific field most relevant to its thesis, genetics. Since species are defined by visible inherited traits, the most important scientific discipline on the question of the origin of species was, is, and always will be, genetics. In the end you can scrap all the other contributing scientific disciplines listed above. Genetics is the only discipline that matters.
Darwin knew very little about genetics because the biological investigation of the cell was in its infancy in his era. He could only see the cell as a moving blob under a microscope. He therefore drew most of his conclusions about inherited traits based on his observations of domestic livestock breeding. Darwin even confessed in 1859 that Our ignorance of the laws of variation is profound (Origin of Species, 1859, p.167). 160 years later that profound genetic ignorance has now disappeared because of rapid advances in the knowledge of how genetics works in the 20th Century, followed by astronomical advances this century since the sequencing of the human genome in 2001.
Until the secrets of the human genome were cracked, the chief criticism of Biblical creation, which is the only alternative scientific model of origins that has seriously challenged the Darwinian model, is that it has failed to make and successfully test significant scientific predictions, and is therefore not science. The scientific method formulates a hypothesis, makes predictions based on that hypothesis, and then rigorously tests them. If the testing validates the hypothesis then a new theory begins to emerge. If contradicted, the hypothesis is scrapped. That key criticism of the Biblical model of origins is no longer scientifically valid. In fact it has now reversed. Creationism’s many genetic predictions are now being repeatedly tested and validated while evolutionary predictions are now failing short of the standards of the scientific method.
This essay, with its emphasis on human genetics due to the abundance of research on our own genome this century, will work its way through 12 scientific predictions that arise from the Biblical model of biological origins, highlighting their recent validation in the scientific literature and their resulting superiority over the evolutionary model of origins. The 12 predictions are as follows:
- Genetic mutations cannot create new species. They only destroy them.
- Bult-in capacity for variation (heterozygosity), is the true cause of speciation.
- Speciation is a rapid process. It did not take millions of years.
- Human DNA can be directly traced back to Noah and his wife.
- Female Mitochondria will contain the echo of three women immediately after Noah’s wife.
- Male Y Chromosomes will also contain the echo of three men immediately after Noah.
- Female mitochondria mutation levels will reflect a recent creation.
- Male Y chromosome mutation levels will also reflect a recent creation.
- The Y chromosome mutation map will be a mirror of recorded history, not pre-history.
- The Y chromosome mutation map will also perfectly reflect 6,000 years of population growth.
- The ever-shrinking level of junk DNA will eventually go to zero.
- Human and chimpanzee DNA is too divergent to share a common ancestor.
Before we get started it is important to clarify two things:
First, the essay will flip back and forth between two types of DNA, so please don’t get them confused. Nuclear DNA is the vast encyclopedia of information contained in the nucleus of every cell and is the blueprint for the construction and maintenance of everything about us. There are around 3.2 billion nucleotides of DNA (think of them as letters of the alphabet; the building blocks of genetic words, sentences, paragraphs, and volumes of instruction) in each cell. Mitochondria DNA is very different. Mitochondria are small energy factories that transform chemical molecules from our food into energy so the cell can function. There are hundreds of them in each cell, and each one contains just 17,000 nucleotides of DNA. The creation model of origins treats the origin of genetic variation in each of these two sources of DNA very differently. The evolutionary model treats all variation in each source as a result of mutations alone.
Second, as you read on make sure you note the difference between vertebrate and invertebrate DNA as you read. The differences in their genetic variation and mutation levels are stark. There is much research to be done on why this is so, but it is early days, and you will read some fascinating recent research on these differences as we go. Now, let’s get started.
1. GENETIC MUTATIONS CANNOT CREATE NEW SPECIES. THEY ONLY DESTROY THEM.
The prime axiom of all Darwinian evolutionary theory, and the bedrock foundation that nearly all else in our post-Christian, Atheistic civilisation stands on is this: Accidental genetic copying mistakes (mutations) can create new and superior genetic information and hence biological function given enough trial and error (natural selection) and enough time.
Mutations are thus the bedrock of evolution. For evolution to be a viable theory there must be a significant percentage of beneficial mutations, mutations that create new and superior genetic information that enhances biological function. In addition, the evolutionary model must eliminate most harmful mutations via natural selection, or the genome will possibly degrade over time toward extinction.
So, where did this idea of abundant positive mutations leading to onward and upward evolution come from? Not long after the rediscovery in the 1930’s of Gregor Mandel’s lost work on genetics, evolutionary scholars such as Ronald Fisher, Sewall Wright and John Haldane (click on “page 174” to get the full comment) rescued evolutionary theory from the dustbin of history by predicting that the ratio of beneficial to harmful mutations in humans is somewhat even, allowing evolution a fighting chance once the magic wand of natural selection eliminated the negative mutations.
Recent studies have shown that both these assumptions are not supported by the evidence. The vast majority of all mutations are tiny point mutations of a single nucleotide, equivalent to a single spelling mistake in a book of 500 million words. These tiny mutations are too small to be adding any useful genetic information, and if harmful they are too small to be eliminated by natural selection. Whether beneficial or harmful, they simply accumulate over hundreds of generations.
In contrast, the creation model of origins predicted the mutational ratio would be completely skewed toward harmful mutations because of the curse placed on creation at the fall of Adam and Eve. Which model of origins was right?
In 1998 the true ratio of beneficial to harmful mutations was finally assessed. It came in somewhere between 1 positive mutation to between 100,000 and 1,000,000 harmful mutations. This is light years from the evolutionary prediction described above. This extreme ratio was again confirmed by Santiago Elena in 1998, by Thomas Bataillon in 2000, and again by Bataillon and Bailey in 2014 (Genetic Entropy p.24-36). Dr Jerry Bergman’s extensive literature review in 2005 found 453,732 mutation hits, but only 186 included the term beneficial mutation, about 4 in every 10,000. Of those 186 references to beneficial mutations, he found not a single one that unambiguously created new, more complex genetic information. This clearly leaves evolutionary theory with too little material to work with, and therefore too little time even with several billion years up its sleeve.
But it gets worse. Any of those 186 hypothetical and exceedingly rare beneficial mutations do not add anything new to genetic complexity. All they can do is jam a regulatory genetic signal in the “on” or “off” position. This is due to the receptor being removed and no longer responding to its signalling molecules. Any perceived benefit is clearly a loss of information from the original genetic instruction. Most jamming mutations create dysfunction but in extremely rare situations they can benefit an organism, such as giving Northern Europeans the ability to tolerate milk in adulthood. These rare and inconsequential benefits do not in any way support the assumption that complex biochemical life has arisen via mutations. Mutations can never be a mechanism that creates a prince from a frog, or an X-man from mere mortals, if given enough time.
On a generation by generation basis this increasing human mutational load is between 80 and 200 new harmful mutations per generation (Nachman and Crowell 2000). At just 80 mutations per generation, and a generous 1:100,000 ratio of beneficial to harmful mutations instead of 1:1,000,000, there would only be a single beneficial human mutation every 1,250 human generations, or around 25,000 years. Most of these beneficial mutations would be tiny point mutations, our single letter copying mistakes mentioned above. To accumulate 100 beneficial mutations would therefore take 2.5 million years, and then they would be scattered randomly among 3.2 billion nucleotides. They would have to be in the exact same location to be of any use. In the meantime they’ve been drowned out by 10,000,000 harmful mutations that would have already pushed us well down the path toward genetic dysfunction and eventually extinction.
There are other types of larger mutations that do have some effects on the entire human genome, and doctors know them well. They are the cause of many genetic diseases in humans. They do not create evolution. Mutations are the cause of around 10,000 genetic diseases and counting. Natural selection thus works to slow down genetic extinction and nothing more. With the recent rise of sophisticated medicine allowing many more people with genetic defects to live to adulthood, leading geneticists hold grave fears for the overall health of the human genome in just a few hundred years from now.
Thus, there is a slowly increasing negative mutational load building inside the human genome, every generation. It’s building in every other species as well. It acts as a slow moving doomsday clock, ticking down to a point, somewhere in the future, when mutational loads will eventually cause the extinction of not only humans, but all life on earth. This dire problem has led human geneticist Alexey Kondrashov to title a 1995 research paper: Contamination of the genome by very slightly deleterious mutations: Why have we not died 100 times over? This heading tells us he is obviously working from a long age evolutionary mindset. Mr Kondrashov, if we are millions of years old as a species, we would have died out long ago!
Conversely, if we move backwards in time and reverse this genetic doomsday clock, we get less and less mutational load. We then end up with a perfect genome for all species sometime in the past, beyond which we cannot go. This scenario fits the creation model perfectly as it predicts that all life was originally created perfect and then began to genetically degenerate (Genesis 3). Thus, the creation model and the iron-clad law of entropy align, and genetic research now confirms that alignment. In layman’s terms the law of entropy says that in a closed system everything moves from order to disorder over time. Everything breaks down. The evolutionary model thus contradicts this law of physics.
2. BUILT-IN CAPACITY FOR VARIATION (HETEROZYGOSITY) IS THE TRUE CAUSE OF SPECIATION.
For over 160 years evolutionary theory has clung doggedly to the belief that all differences between any two life forms are only due to mutational change and natural selection over time. There is no other cause permissible. But consider the following: Assume the mutation rate for humans is around 80 per generation, and the evolutionary model says we emerged out of Africa around 200,000 years ago, and assume we had a 20 year reproductive cycle. This brings us to an evolutionary prediction of around 800,000 single nucleotide mutational differences between people (200,000 divided by 20, times 80). However, studies have confirmed that there are over 4 million nucleotide differences within the humane gnome between people. This is fivefold higher than predicted by the evolutionary model. The nuclear DNA difference between chimpanzees and humans is predicted by the evolutionary model to be between 8 and 13 million nucleotides. Instead, it is around 25 million. Why?
It turns out that mutations are not the only potential cause of genetic change within an organism. In fact they are one of the least significant sources, and as we saw above, they are almost universally negative, so it’s a good thing they are insignificant. The main source of genetic difference between humans is genetic recombination of male and female chromosomes. We see it visually in our children’s mixture of parental features. But that’s not all. Our DNA has 23 chromosomes, but we have two copies of each of these chromosomes, so 46 in total. The constant recombining of these duplicate chromosomes leads to individuals where, for example copy 1 and copy 2 of chromosomes 15 or 21 or 6 etc, are not quite identical. This difference is called Heterozygosity.
To illustrate, let’s consider Darwin’s use of domestic breeds as his prime argument. The vast amount of variation between breeds of dogs, ducks, cats, and cattle has never been claimed by anyone to be caused by mutational change. All parties acknowledge the potential for the amazing variation we see in domestic animal breeds was always there hiding in the original DNA. Artificial breeding just teased it out. Everyone also acknowledges there are natural limits to this variation. So Darwin’s number one example of evolution had nothing to do with evolution, just in-built potential for variation up to a genetic limit, which is exactly what the modern creation model predicts.
Studies have now shown that the more genetically distant two parents are, the more heterozygous will be the two copies of their same chromosomes, and this will be repeated in their offspring. This results in a higher speed of physical change in descending generations of offspring (Replacing Darwin p.218). Once again, we see this when people from different races have children. In addition, because more of the 46 chromosomes are different, highly heterozygous individuals also produce offspring with higher levels of mutational damage simply because of the higher level of mismatch in their chromosomes. However, this mutational acceleration is a far smaller cause of genetic change than heterozygosity, and it is almost universally harmful to the organism.
Now let’s switch from looking at differences within a species to differences between species. Evolutionists use pre-existing heterozygosity to explain that 50% excess genetic variance between humans and chimpanzees mentioned above that is not predicted by their model. But they then jump back to their evolutionary doctrine by claiming this pre-existing heterozygosity is a product of prior mutational change deeper in evolutionary time. Now let’s take that logic to its natural conclusion: If evolutionists can use the heterozygosity argument at the human/chimp level, then creationists can use it too, and then take it further and further back to its beginning point; heterozygosity in the creation of a perfect genome that is now breaking down. The creation model says all organisms were designed with in-built genetic heterozygosity at the point of origin. These pre-designed levels of heterozygosity would have allowed for very fast genetic change over just a few generations.
The Creation model therefore does not rely on mutations, a slow, cumbersome, and inefficient mechanism that has now been proved to be utterly destructive to our survival. In contrast, heterozygosity allows for very fast speciation. A point we will now explore.
3. SPECIATION IS A RAPID PROCESS. IT DID NOT TAKE MILLIONS OF YEARS.
The classification system for all life forms begins at the species level and then goes up through genus, family, order, class, phylum, and kingdom, to domain. We all learnt about it in high school biology. The evolutionary model predicts that all differences at all these levels in all life forms are the product of unguided mutations and natural selection over billions of years.
Alternatively, the creation model predicts that at the time of design and creation 6,000 years ago, the exact level at which this design and creation took place in vertebrates was not at the species, or even the genus level but rather at the family level. This level of creation is called kinds in scripture (Genesis 7:1-4). The creation model also says that for invertebrates, their design and creation was at the genus level. The creation model also predicts that in the 6,000 years since creation we have seen the rise of all the species which we see today from those original kinds. There are around 69,000 vertebrate species but only 1,123 vertebrate families, so species are over 98% of that total.
How could so many species arise in such a short period of time I hear you ask? The answer of course goes back to the magic of heterozygosity, but first let’s circle back to Darwin’s analogy of livestock breeds.
The vast array of often bizarre breeds in all forms of domestic and pet animals are the product of rapid change over just a few thousand years, and there are far more breeds than wild species for each of these domesticated animals. For example, there are many hundreds of horse breeds, but only around 80 extinct species, and just 7 living wild species in the equine family: Imperial zebras, mountain zebras, plains zebras, the kiang, the African wild ass, przewalski’s horse, and the onager. For context, there are between 10 and 28 million nucleotide differences between the wild species just mentioned, and around 6 million between our domestic horse breeds.
Darwin assumed the process of biological change would be much slower in wild species both because it was unguided and because of his prior commitment to deep time, even though the evidence from the rapid breeding of domesticated animals told him otherwise. If evidence from the farm was good enough to be used as proof for evolutionary speciation, then Darwin was not being intellectually consistent. Why should 87 species of equids take 40 million years when hundreds of breeds of horses took a few thousand years? That’s a ratio of 4,000:1!
Because of their prior commitment to deep time, evolutionists do not expect to find evidence of rapid speciation in real time, so they haven’t been looking. No one actually knows the true speed of speciation because until 50 years ago no one had the scientific means to do the research over several generations, long distance travel was uncommon, human lifespans were short, no one was looking for new species, and education levels were relatively low. Having said that, an example of rapid speciation has recently been recorded among Darwin’s famous Galapagos finches no less (Sangeet Lamichhaney et al., Rapid Hybrid Speciation in Darwin’s Finches.). What the researchers thought would take hundreds of generations took only three!
The magic of heterozygosity, the front-loading of DNA differences between two twin chromosomes of the same number at the original creation, is the cause of rapid speciation. It did it for domestic animals, and it did it for wild animals just as fast. Imagine the possibilities if all 46 chromosomes were heterozygous at creation instead of one or two? The possibilities are almost endless. A strongly heterozygous small number of breeding pairs at the original creation would have produced offspring with huge nuclear DNA variance. If several of these highly heterozygous individuals migrated to isolated areas after a small number of breeding cycles, they would have been cut off from the wider gene pool, thus accelerating and locking in any physical changes in traits through the loss of genetic material a.k.a. natural selection. For example, white bears living in snow country losing the DNA for black fur.
In summary, all we need in order to form a new species is a small and highly heterozygous group of animals that become isolated from the original population early in history. Physical inherited change will be rapid while at the same time inbreeding will be inevitable, locking in those physical changes. This one-two punch will knock out much of the prior heterozygosity. As the population grows (and populations always grow exponentially until resource constraints kick in) its level of heterozygosity continually diminishes, locking in the physical distinctions from the original population. Because of the in-built heterozygosity at creation, and the rapid migration over the planet after the flood, we have the perfect ingredients needed for rapid speciation without the need for millions of years and mutational change, which as we saw in the very first section of this essay, cannot produce any meaningful new genetic material anyway.
When humans reverse this natural process with closely related but distinct species we find that many separate species can still actually breed together, resulting in some amusing new hybrids such as a liger (a cross between a lion and a tiger), a zebroid (a cross between a zebra and a horse), and a wholphin (a cross between a killer whale and a dolphin). This is yet more proof that these species had a recent common ancestor further back in history.
{kind=link}
Instead of asking why so many new have come about in just 6,000 years, we should actually be asking why there are not more. This is predicted in the creation model of origins and is exactly what we see in the fossil record. Rapid extinction is the flip side of the rapid speciation process. In fact it is the rule rather than the exception. There are 16,000 species of mammals in the fossil record, but only 5,400 alive today. Over 550 mammal families exist in the fossil record but only 150 are still with us. So much of the original creation is gone forever because of the curse. As previously mentioned, there are 80 extinct equid species and only 7 living species (Replacing Darwin p.275). Many new species in the past have been so inbred that they were unsuited to their new environments and could not cope. Mutations did not build them, they destroyed them. This is why the dinosaurs died out. This is why the Neanderthals died out. Ongoing speciation and increasing extinction will continue until the end of time. It’s the natural order.
4. HUMAN DNA CAN BE DIRECTLY TRACED BACK TO NOAH AND HIS WIFE
If Noah and his wife were real people then surely there is a clue to their existence in our genes. This has always been a key prediction of the creation model, and it is now confirmed. Scientists have long known that the Y chromosome in men and the mitochondria in women are passed down with almost zero influence from the opposite sex. This means they do not mix and match at fertilisation but are passed down basically unchanged from father to son and from mother to daughter. In the process they slowly accumulate mutations without any natural selection eliminating these mutations. In this regard they are very different from normal DNA. This unique feature allows us to track ancestry.
A woman’s egg has thousands of mitochondria while the male sperm has almost none. This means almost 100% of all mitochondria are inherited from mum, grandma, great grandma etc. If two seemingly unrelated people share the same mutation in their mitochondria this means they once shared a common female ancestor. The mapping of these mutations can now be undertaken across whole populations, and scientists call these mutational maps haplogroups. You can watch a good video introduction to haplogroups at this link. Haplogroup mapping has allowed scientists to track female mitochondria mutations right back to our beginnings, and it turns out the full mitochondria haplogroup map points back to a single woman somewhere in our ancient past, whom secular scientists euphemistically call mitochondria Eve. The evolutionary explanation for this amazing genetic discovery is that this woman was a bottleneck in the ongoing evolution of humanity. The creation model says it was not actually Eve, but Noah’s wife as she was the most recent genetic bottleneck for humanity.
In men it is only the Y chromosome that differentiates them from women. Men have an XY chromosome while women are XX. All chromosomes are nearly identical and are mixed at fertilization, except for the Y chromosome. The Y chromosome is handed down from father to son unchanged, so the male Y chromosome also creates a mutational trail that leads back to a single male in ancient history, whom secular scientists have once again euphemistically called Y chromosome Adam, or Most Recent Common Ancestor (Y-MRCA). Evolutionists explain the existence of this Y chromosome Adam by suggesting this man lived up to 200,000- 300,000 years ago and was once again just the most recent bottleneck in the ongoing human evolutionary tree. The creation model tells us this man was Noah, since he was the most recent bottleneck in male Y chromosome biology. Y chromosome Adam should be renamed Y chromosome Noah!
Regardless of the secular explanations, the fact remains that the creation model of origins predicted that human DNA would reveal a historical bottleneck as small as a single man and a single woman. What are the chances of finding two single-person bottleneck ancestors in human ancestry by chance? The validation of this key creation hypothesis strengthens the theory significantly.
5. FEMALE MITOCHONDRIA CONTAIN THE ECHO OF 3 WOMEN IMMEDIATELY AFTER NOAH’S WIFE.
Now let’s move on to the children of Noah. If three historical sons of Noah came off the ark with three historical wives, then this allows the creation model of origins to make another prediction: We should be able to find the echo of this historical fact in female mitochondria. And this is exactly what we find. As we saw above, a woman’s mitochondria DNA mutations are not subject to chromosomal recombination. The slowly increasing mitochondria mutational load is therefore a brilliant and simple way to track genetic ancestry. If you share a mutation, you share an ancestor. You can see almost the entire historical female mitochondria haplogroup at this link. What is immediately visible is that the mitochondria mutational tree cascades out from three women (green arrows). You are actually looking at the genetic footprint of the three wives of Shem, Ham, and Japheth! The lines connecting the three initial women are pre-flood mutations and is therefore our only genetic glimpse of life before the flood.
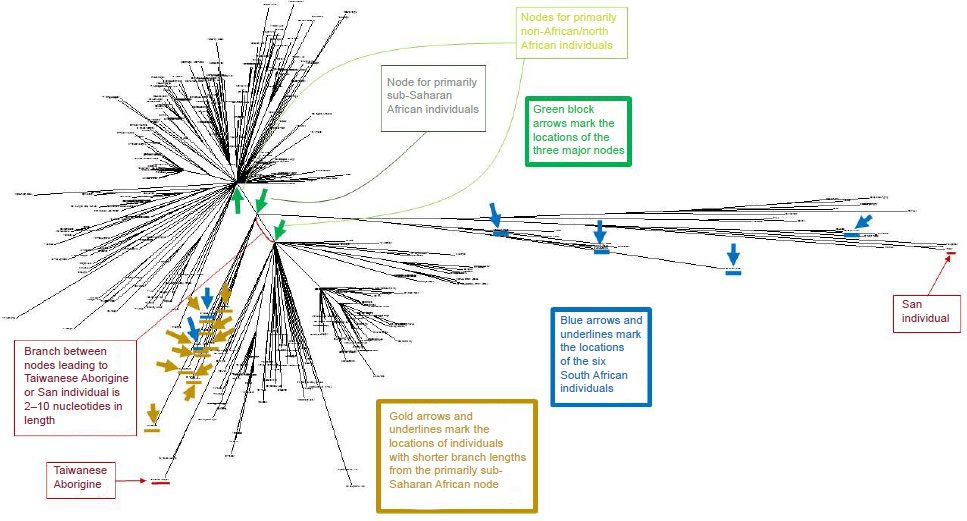{kind=link}
The longest branches in the haplogroup tree correspond to those individuals with the most mutational differences to others, which is African women. African women come in with around 123 mitochondria mutational differences, almost twice as many as other women. If women in Africa have been reproducing at a younger age than in other parts of the world this difference is fully accounted for. Alternatively, there may be a slightly faster mutation rate in African women, which is a prediction waiting to be tested. The shape of the haplogroup tree represents globally significant migrations, famines, wars, empires, and plagues. All that by just looking at mitochondria mutation links! Once a mutation occurs it creates a new branch in the tree because the mutation will be unique to that woman and her descendants. You are literally looking at the rise and fall of civilisations throughout history via a genetic map.
6. Y CHROMOSOMES WILL ALSO POINT BACK TO 3 MEN IMMEDIATELY AFTER Y NOAH
The creation model of biological origins also predicts that there will be evidence of exactly three descendants from Y chromosome Adam (Noah) to match the three descendants of mitochondria Eve (Noah’s wife) mentioned above, confirming the Biblical account of Noah’s three sons, Shem, Ham, and Japheth (Genesis 10).
Many genetic studies have now been conducted on Y chromosome mutational differences between men from all around the world. Scientists have mapped these unique differences into groups that share a common ancestor. As we just learned, these groups are called haplogroups, which look like a giant, complicated family tree, which they are.
What is fascinating is that the Y chromosome haplogroup tree keeps joining as you go back through history until it reaches exactly three separate branches immediately after the original Y chromosome Adam (Noah), and from there it branches rapidly to all men on earth as people dispersed around the planet. If you look carefully at this link, you will see the original three original men at the very right of the link, and top of the diagram.
{kind=link}
This recent discovery is yet another confirmation of the creation model of origins. Secular scientists will acknowledge this three-part split but place it hundreds of thousands of years earlier based on their understanding of human origins. This assumption is not based on genetics, but a prior commitment to a long-ages geological framework, without which evolution cannot be a viable theory. Once again we see the human Y chromosome haplogroup map corresponding to Biblical history.
7. FEMALE MITOCHONDRIA MUTATIONS NUMBERS WILL REFELCT A RECENT CREATION.
If the total number of mutations in female mitochondria were known, along with the rate of new mutations per generation, we could divide one by the other and have at our fingertips an objective genetic clock that could measure how old we were as a species. This would supersede the need for inferior external clocks such as carbon and radiometric dating. It’s a simple clock too: If we as a species are millions of years old, there would be many mutational copying mistakes. If our species has a recent origin, there would be few. Using evolution’s own assumption that what we measure in the present has been consistent in the past, we will also assume this genetic clock has always ticked at the same rate we observe today.
The results are now in. The average female mitochondria mutation rate is one new mutation every 3 to 7 generations of women, or one per 75 to 175 years assuming 20 years per generation. On the other side of the equation, the total number of mitochondria mutations in the average mitochondria is around 70-80 for the average woman (however, as we noted above, African women average an extra 77 mutations). Multiplying 80 mutations by 3 generations per mutation, by 20 years, gives us exactly 4,500 years. Multiplying 80 mutations by the maximum 7 generations by, 20 years still only gives us 11,000 years. These figures are all millions of years outside the evolutionary timeframe.
If we were a product of evolution, and we separated from chimps at 4.5 million years ago, there should be between 25,000 and 60,000 mutations inside the average mitochondria. This is an impossible number since there are only 17,000 nucleotides of DNA in any mitochondria.
By the way, the mitochondria mutation levels for chimpanzees came in at around the same tiny level as humans. The same goes for the Neanderthals, whom the evolutionary model claims diverged from us between 400,000 and 700,000 years ago. The evolutionary model predicts between 1,660 and 9,200 mitochondria mutational differences between Neanderthals and modern humans. However, there are just 213. The creation model predicts that these 213 mutations are not necessarily because of time but that the Neanderthal DNA mutated too quickly after the flood and they as a group died out faster than we are dying out. This prediction is still to be tested.
Evolutionists also claim Africans are the oldest humans because of their extra mutation levels, and the rest of us started moving out of Africa some 200,000 years ago. If this was the case, there would be between 477 and 2,600 mitochondria mutational differences between African women and the rest of humanity. As we saw above, there are just 77. As mentioned previously, this difference is probably not because of time but because of either a consistent difference in average marriage ages of women in Africa compared to the rest of the world over the centuries, or a slightly faster mutation rate.
Mitochondria Mutation Levels:
| Group | Evolutionary Prediction | Creation Prediction | Result |
| Humans / Chimps | 25,000 – 60,000 | 450 – 2600 | 1,483 |
| Moderns / Neanderthals | 950 – 9,200 | 14 – 79 | 213 |
| Africans / Non Africans | 477 – 2,600 | 14 – 79 | 77 |
The mitochondria mutational evidence continues to mount in other species as well, and it is all in favour of the creation model. Below are results from mitochondria studies in four invertebrate species.
Mitochondria Mutation Levels
| Species | Evolutionary Prediction | Creation Prediction | Result |
| Roundworm | 45,000 – 9,000,000 | 15 – 3,000 | 1,678 – 1,801 |
| Fruit Fly | 45,000 – 10,000,000 | 5 – 1,200 | 865 – 1,778 |
| Water Flea | 22,000 – 6,000,000 | 18 – 5,000 | 3,173 – 4,675 |
| Baker’s Yeast | 2,000,000 – 200,000,000 | 840 – 80,000 | 5,042 – 19,670 |
The creation model predicts that the results outlined in the table above will be consistent for the millions of other non-vertebrate species yet to be analysed, and those results are starting to filter in now. One such study by Stoeckle and Thaler even tries to solve the obvious evolutionary dilemma by suggesting that all animal species came through a single mitochondria ancestral bottleneck within the last 200,000 years in a similar fashion to mitochondria Eve and Y chromosome Adam. They even demand the evolutionary community give a better explanation! Evolution’s response will be predictable, they will once again wave the magic wand of natural selection. However, as we saw in Sections One and Two, natural selection is more like a spiked club than a magic wand.
When it comes to vertebrate species there are very few mitochondria count studies apart from humans. The results of three studies on mice, chickens, and adelie penguins disagree with both the evolutionary and creation models. Both models are out by orders of magnitude. It is early days and these three are the only studies available. In the initial research on human mitochondria mutation count measurements in the 1990’s also fluctuated wildly, but then coalesced around a reliable figure. Many more tests need to be done on vertebrates. This in no way diminishes the significance of the mitochondria results in both humans and invertebrate studies, it just means we are still in the initial phase of this learning curve.
8. MALE Y CHROMOSOME MUTATIONS LEVELS WILL ALSO REFLECT A RECENT CREATION
As we’ve seen already, every Y chromosome in every man has slowly been accumulating mutations in the same way as female mitochondria. Therefore if we know how many mutations in total there are in an average Y chromosome, and how many mutations are accumulating per generation, and divide the number by the rate, it will also tell us how many generations have passed since we began. The Y chromosome mutation level and rate is therefore another genetic clock going tick-tock inside our male bodies.
The Y chromosome mutation rates have recently been published in four studies that measured father/son pairs. The first two, (Xue et al 2009, and Helgason et al 2015), were under considerable budget and time constraints, so they filtered out difficult-to-read parts of the Y chromosome. Their incomplete studies showed a mutation rate very slow and in conformity with the established belief that humanity emerged around 200,000 years ago.
But then another higher quality study (Karmin et al 2015) came up with big surprise, a raw figure between 10 and 17 times faster than the first two studies. Sadly and not surprisingly, this study then filtered this shock result so that it conformed with consensus evolutionary beliefs about the 200,000 year human timescale. The final high quality study (Maretty et al 2017) compared the DNA sequences between 62 parent/child pairs that included several father/son pairs. Suspiciously, they published results on many parent/child DNA differences except the father/son chromosome data, of which there were 17 pairs. Dr Nathaniel Jeanson (Traced. 2022, Replacing Darwin. 2017) has examined the raw data for the study and found it confirmed Karmin’s 2015 study that Y chromosome mutation rates were running way too fast, at around three per generation, many times faster than the original researchers were comfortable with.
Because of Dr Jeanson’s detailed analysis of these two latter studies we can now confidently predict that future studies will confirm the findings that the total number of Y chromosome mutations is an average of around 450 per male alive on the planet now, and the Y chromosome mutation rate per generation is a very rapid 3 per generation (Replacing Darwin p.212). If we divide 450 mutations by 3 per generation we can find the time when the Y chromosome stopwatch started ticking. This works out at roughly 150 generations ago (450 divided by 3), or about 4,500 years if we take 30 years as a generation for men as they were historically older than women when first married. This just happens to be the timeframe from the great flood event of Noah to the present.
Tweak it any way you like, and it still comes out in the ballpark of a recent creation. That’s a big miss and a huge embarrassment to the evolutionary timescale, so they claim once again that good old natural selection has filtered out most of the ancient, accumulated mutations. However, as we now know, nearly all mutations are tiny point mutations that can never be filtered out by natural selection which only takes place at the person level, at reproduction, not at the DNA level. Natural selection can therefore only filter out massive genetic mistakes such as Down Syndrome, but never a tiny mutation.
The Y chromosome is therefore giving us another new clock with which to measure human history on earth. A clock superior to, and in direct conflict with the standard geological model with its three inbuilt assumptions of zero contamination over millions of years, constancy of decay for millions of years, and zero daughter isotopes at the time of formation of origin for radiometric dating systems. The fact that the Y chromosome clock confirms the measurements of the mitochondria clock is highly significant, right up there with the discovery of a single man and woman in our DNA ancestry.
9. THE Y CHROMOME MAP WILL MIRROR RECORDED HISTORY, NOT PREHISTORY.
Dr Nathaniel Jeanson’s book, Traced, (which is neatly summarized in this link), has given the world a completely new way of looking at our common human ancestry. As we saw above, the evolutionary timescale does not agree with the mutation levels and mutation rates for both Y chromosomes and mitochondria. These two clocks allow us to confidently shift the timescale of the human Y chromosome haplogroup map mentioned previously from 200,000 years to a 4,500 year timeframe, the genetic bottleneck of the great flood, a task Dr Jeanson has done for us.
This shifted timescale allows us to make another clear creation model prediction: That the male Y chromosome haplogroup map introduced in Section Six does not record human history going back 200,000 years but is a clear reflection of recent and recorded human history such as verified historic migrations, plagues, wars, civilizational collapse, disease, and the rise of today’s language families, right down to regional and local levels. This level of historical detail in the Y chromosome haplogroup map is in addition to the three sons of Noah at the very top of the map that we previously talked about. All these ebbs and flows in human population are reflected in the physical shape and size of the male haplogroup map. This confirmed prediction is the ultimate confirmation of the accuracy of the 4,500 year timeframe for human history since the flood. For far more detail than the few paragraphs below, I urge you to read Dr Nathaniel Jeanson’s excellent book. What you read below is just the tip of the iceberg:
If we break the Y chromosome haplogroups down to regional levels, we find Dr Jeanson’s assumption of a recent human origin accounts for the E1b1b haplogroup emerging from the Middle East and moving into North Africa at the same time as the Arab conquests of that region. It also accounts for the rapid spread of the E1b1a haplogroup across sub-Saharan Africa from around 1400 AD as people in that region fled the slave traders. In fact, haplogroup histories are telling us that humanity did not come out of Africa at all but moved into Africa via Egypt thousands of years earlier. This pattern agrees with the Tower of Babel thesis. Jeanson’s timescale adjustment also accurately accounts for the emergence in Western Europe of Central Asian DNA from the same time the Ottoman Turks(haplogroup J2) overran Constantinople in 1453 and then pushed all the way to Vienna. This then pushed East Europeans (R1b-U106, R1b-S116 and R1b-U152) into western Europe from the time of the Ottoman invasion. The list could go on and on. Nearly everyone with a Western European ancestry now has both East European and Central Asian DNA!
Dr Jeanson’s timeframe adjustment also accurately maps the movement of the Magyars (haplogroups R1b-M73 to R1b-M269) from central Asia to Europe just over a thousand years ago. These became the Hungarians we know today with their unique language that is linked to Asia, not Europe. Jeanson’s adjusted haplogroup map also accurately traces the movement of the original cousins of the Hungarians, haplogroup R1a, from Central Asia down into the lands south of the Himalayas with the arrival of the Mughals in the Indian Subcontinent. Recorded imperial history matching genetic history.
The haplogroup map tells us the Melanesians, the Polynesians, and the Australian Aboriginals (haplogroups C, K, M, S), came to their new homes in the millennium before Christ, that the Polynesians have much in common with Siberians (haplogroup C), that Polynesian ancestry can also be traced to the original inhabitants of the Philippines, and that there were two waves of migration into Australia (haplogroups C and S), also indicating a minor connection to Siberia. I could go on for hours!
A neat confirmation of the accuracy of this revised haplogroup timeframe comes from the natural organisation of the many unique human language families. Many languages come from common ancestors and these branch languages can be grouped into large language families that have distinct geographical and historic footprints. These language family footprints are confirming the distribution and progress of the recent movement of haplogroups across the earth. One example among many is the inclusion of the north Indians and most Europeans in the Indo-European language family. We share a common ancestor, and this is why the connections between Sanskrit and ancient Greek is so strong.
10. THE Y CHROMOSOME MAP WILL PERFECTLY REFLECT 6,000 YEARS OF POPULATION GROWTH.
An offshoot to the research just mentioned lies in the global human population growth profile. The written record of human population growth throughout history goes back to around 1,000 BC. That record is basically a chart of a steady and slow increase that was slowed occasionally by a few dips. Steady global growth then gives way to exponential growth around 1600 AD as we began to master fossil fuel energy sources. Only if we start from 4,500 years ago, instead of the hundreds of thousands of years traditionally assigned to human haplogroup history, do we get an exact fit between known recorded population growth and the shape of the Y chromosome haplogroup map. There is a 90% overlap in the multiplication of humanity and the multiplication of haplogroup mutational branches only if we begin from that starting point. If we evolved 200,000 years ago there is no overlap at all between the haplogroup mutational map and known population growth. If we assume humanity came out of Africa there is only a 60% fit. The roots and spreading branches of the Y chromosome haplogroup map are not recording changes in population size from 200,000 years ago, but changes in population size from the recent past.
11. THE EVER-SHRINKING LEVEL OF “JUNK DNA” WILL EVENTUALLY GO TO ZERO.
Evolutionary theory insists that billions of years of mutations will inevitably lead to unusable pieces of junk genetic code floating around in our DNA. In opposition to this prediction, a core premise of the creation model is that all our genetic code is fully functional and essential. When the human genome was first fully sequenced in 2001, evolutionists trumpeted the fact that 99% of our genome that was non-protein coding DNA. This meant the 99% of our DNA did not build stuff inside the cell so was probably evolutionary junk. The genome had not been fully tested at that point so this was purely a theoretical assumption. Research into that 99% of our DNA then took off in a big way and in 2012 the Encyclopedia of DNA Elements (ENCODE) project published their research on the human genome. They found that at least 80% of our DNA was functional. These results were preliminary but are on the right track.
Many new discoveries in recent years are now pushing this level of functionality even higher. The leader of the ENCODE project, Ewan Birney, is predicting that the human genome will soon prove to be 100% functional. Needless to say, the traditional Darwinian evolutionists outside the biomedical genetics community are outraged that the data is not supporting their evolutionary model. It is only a matter of time now before science will discover functionality in the remaining 20% of our DNA. If this is the case for humans, it will also be the case for all species, confirming the creation prediction.
For other simple species recently tested we have found that nearly 70% of roundworm pseudogenes, the technical term for non-protein coding segments of DNA, show biochemical signs of functionality. For fruit flies some 89% of the entire genome now shows preliminary evidence for function. Given this trajectory it looks likely future research will all flow in the direction of the creation model of origins.
12. HUMAN AND CHIMPANZEE DNA IS TOO DIVERGENT TO SHARE A COMMON ANCESTOR.
Now let’s circle back to chimpanzees for one final time. If evolution was true, and mutations are the only way genetic material can change over time, and there are approximately 80 new mutations each generation, and we’ve been separated from chimps for between 4 and 6 million years, then there should be between 18 and 80 million DNA differences between chimps and humans out of 3.2 billion nucleotides, or between 1-3% genetic differentiation. This is a figure you have no doubt heard from time to time in the popular media. However, the results of recent studies show a 12% difference, or 400 million DNA letters out of around 3.2 billion. This is an impossible number to generate via mutations alone.
Because of this predictive failure of the evolutionary model, some researchers quickly changed their original human-chimp ancestral departure point from around 6 million years back to between 13-17 million years. That’s called retrofitting prediction to fact, and it throws out the entire evolutionary story as it pushes everything further back in time. We share 88% of our DNA with chimpanzees simply because we were designed and built in a similar style, with arms and legs etc. We also share 50% of our DNA with yeast simply because it shares the same cell mechanisms.
The process of change from chimpanzee to human is vastly complex. Evolutionary genetic change is not simply about a scattered beneficial mutation here or there, but accumulating vast numbers of beneficial mutations in a single location that will, in time, create a completely new biological function superior to existing functions. Dawkins calls it climbing Mt Improbable, but by now you can see that it is actually climbing the sheer cliffs of Mt Impossible. Part of the problem is that the organism must pass through a genetic fitness valley as the uselessness of the biology of its new genetic material makes it less genetically fit and more prone to elimination in the struggle to survive. For a more in-depth investigation into why it is impossible to create a single new gene, let alone humans from chimpanzees click on this link.
CONCLUSION
Evolution is a theory in crisis. Its prime axiom is faltering under the continual barrage of failed predictions and multiplying contradictions. Its holy grail of beneficial mutations turned out to be a deeply entrenched myth. Mutations are sending us extinct, and all natural selection can do is slow down that inexorable process of extinction. Evolution’s need for deep time also turned out to be a myth, species can form very quickly. Evolution’s confidence in mutations as the prime cause of speciation is also disintegrating as we learn more about heterozygosity, and Darwin’s many references to domestic animal breeds was actually an argument for in-built creationist heterozygosity, not evolution.
In stark contrast, the creation model of origins correctly predicted the universally destructive nature of mutations, rapid speciation, a very recent human genetic bottleneck of a single man and a single woman, then three men and three women immediately afterward. It correctly predicted a tiny number of mitochondria and Y chromosome mutations embedded in the human genome pointing to a recent perfect human genome. It correctly predicted the perfect marriage of the Y chromosome haplogroup map with recorded world history, as well as its perfect match with recorded human population growth. It also is on track to complete its prediction that all DNA is functional.
The pieces of the human origins puzzle are finally coming together. We are a created being, something every culture understood instinctively until 160 years ago when we had the collective audacity and wilful pride to declare we made ourselves. The genetic proof of our exquisite design and function are now being slowly unveiled to that wilful world. Hopefully the gradual revealing of this truth over the next 160 years, starting from around the year 2000 when we finally deciphered the human genome, will be strongly resisted by an entire civilisation that has built its intellectual and moral compass around a lie. Yet in the end they will not be able to hide from brute facts. The eventual acceptance of the truth by our civilisation will the defining achievement of science in the next century as we gradually unwind the lie we have collectively loved.
We will have come back to our roots, back to common sense, back to our creator.