
The evolutionary prime axiom says that every nucleotide, codon, protein, gene, chromosome, protein molecule, cell, tissue, organ, individual and entire species is the result of random, unguided mutations. But what exactly are mutations? Mutations are accidental copying errors in the DNA and RNA transcription (copying) processes that lead to structural change. There are many different types of mutations. The most common one is called a point mutation which occurs when the processes of cellular transcription to RNA make a single spelling mistake. Other types of mutations include accidental deletions, insertions, duplications, translocations, inversions, conversions and mitochondrial mutations. Contrary to common sense, evolutionary theory assumes these copying mistakes can add genetic complexity to all DNA, turning simple life into sophisticated life. It is the same as assuming copying mistakes in a manual for little red wagons can, over millions of years, produce a space shuttle.
1. The 100,000:1 Mutational Ratio
Mutations are very dangerous for any organism and are a major curse in modern medicine. So, in the 1930’s, in an effort to save evolutionary theory from the rediscovery of Mendelian inheritance and a new-found awareness of mutational damage, influential geneticists such as Ronald Fisher, Sewall Wright and John Haldane suggested mutations must fall into a typical bell curve, with half deleterious and half beneficial. Because they believed in the prime axiom, they assumed the deleterious half were then eliminated via natural selection, while the beneficial mutations caused ever upward evolutionary advancement.
Fisher’s knowledge of genetics was rudimentary compared to our own. We now know the real ratio of deleterious to beneficial mutations. It is definitely not 50/50. It is hideously skewed toward the deleterious side. By 1979 the famous geneticist Mootoo Kimura no longer even bothered to record any beneficial mutations on graphs in his research papers, showing mutations to be universally deleterious and exponentially more common as their effect diminishes. In 1998 Phillip Gerrish and Richard Lenski estimated the ratio of beneficial mutations to deleterious mutations to be about 1,000,000:1. That figure is not a typo. They believed there was only one single beneficial mutation to about one million deleterious mutations. This is much closer to common sense and light years away from the assumptions held by Ronald Fisher.
A year 2000 research paper by Thomas Bataillon stated “…the net effect of spontaneous mutation is indeed deleterious. Mean decline of the fitness components of MA lines ranged from 0.1% to 1–2% per generation…Fitness erosion seems to be the rule over a broad range of organisms.” Bataillon believed the ratio of deleterious to beneficial mutations so low as to thwart any actual measurement.
Bataillon and Bailey in 2014 stated that “Direct experimental evidence confirms predictions on the DFE (distribution of fitness effects) of beneficial mutations and favours distributions that are roughly exponential but bounded on the right. (In other words, they agree with Mootoo Kimura. They go on to say) “The conventional assumption…is that populations are very close to their fitness optimum, and so beneficial mutations are exceedingly rare and can be safely ignored. Thus, in most cases, assumptions of the models do not allow for beneficial fitness effects to be estimated at all.”
We can therefore safely estimate the ratio of deleterious to beneficial mutations at somewhere between 1,000:1 up to 1,000,000:1, and closer to the second figure. For arguments sake let’s be generous to evolution and put it at 100,000:1. Unfortunately you will not find this hard science on popular evolutionary websites. Ronald Fisher’s lovely bell curve turned out to be a vastly one-sided, with an exponentially larger number of mutations as they diminish there impact on heritability and effects on the organism. Most of these mutations are the smaller point mutations and are therefore skewed to the central near-neutral zone no-selection zone you see below. This is because the smaller the mutation, the more easily it will slip through the cell’s thirty-plus layers of proof-reading systems.
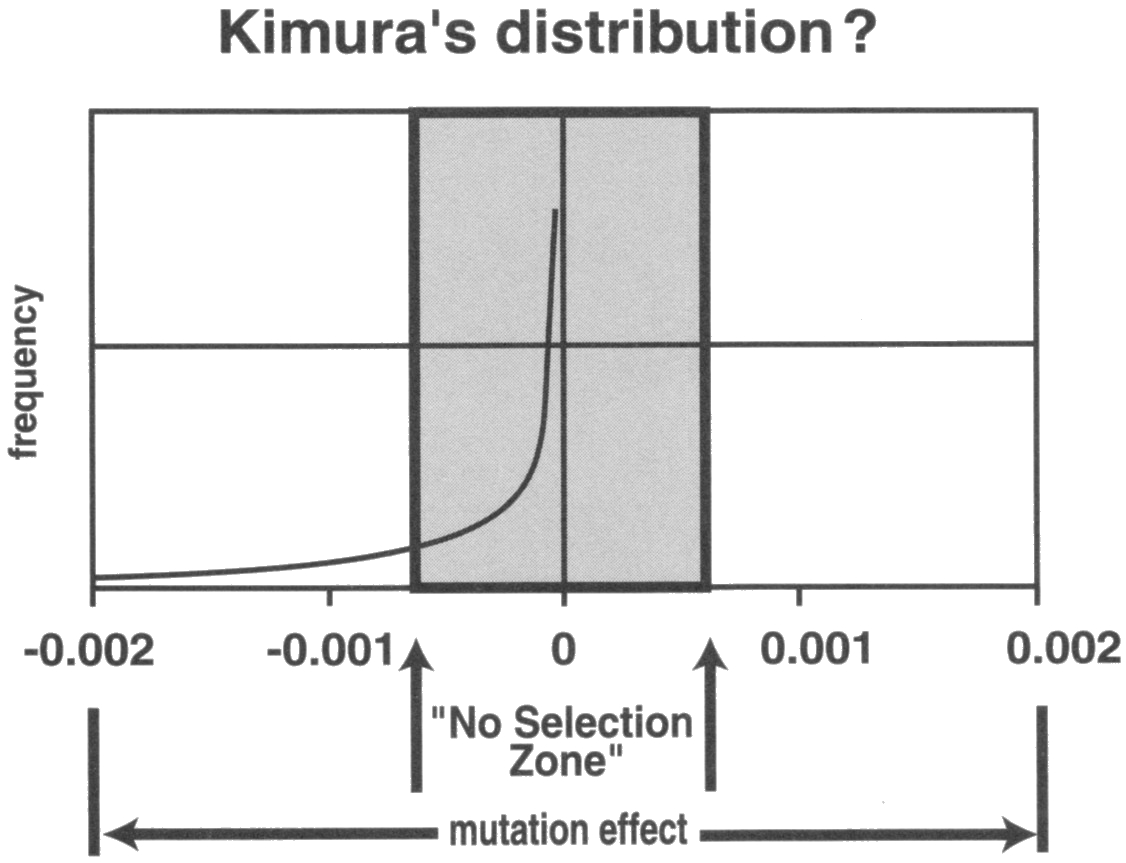
This hundred 100,000:1 raises a profound problem for evolutionary theory. It leaves species development with both too little material to work with and too little time, even given billions of years.
Let’s go through the figures. Here are the assumptions:
- 1. 300 mutations per individual (more on that figure later), per generation
- 2. A 6 year reproductive cycle in chimpanzees stretching to a 20 year cycle in early humans (so an average of 10 years)
- 3. A ratio of 100,000 deleterious mutations to every beneficial mutation
Given these realistic assumpytions it would take an average of 3,000 years for the very first mutation to appear, at any random location. Most of the time this mutation will be then lost due to drift, noise, epistasis, linkage blocks, epigenetics and a host of other factors discussed further on in this essay. This pushes our time frame out by a factor of ten. It’s actually a factor of 100 but we will once again be generous to evolutionary theory. So then we have to wait 30,000 years for the second mutation. But the second mutation could be anywhere, no one and nothing determines where in the 3 billion-nucleotide genome it will occur. So, in 60,000 years you could get your second mutation at opposite ends of the genome! But you want them to occur next to each other so that meaningful change can take place. Don’t hold your breath, as you are looking at a 3-billion-to-one chance of this happening every 60,000 years! In the meantime our pre-human primate has accumulated and locked in some 1.8 million deleterious mutations, and it doesn’t matter if they are in random locations. They will all cause damage.
To get around this problem, many human geneticists have arbitrarily assumed unlimited time when constructing their mathematical models. They have also assumed nearly all mutations have zero negative effect on biological fitness, the perfect heritability of all beneficial mutations, mutual cancellation of deleterious mutations via epistasis, and artificial instead of natural selection. But clearly these assumptions do not reflect reality. Yet they are constantly included in research papers to get around the intractable problem of this mutational ratio. They, and the entire world of biological science, are stuck with a deleterious mutation ratio that has destroyed the prime axiom, and is destroying the credibility of evolutionary theory.
But it gets worse…
2. Beneficial Mutations Don’t Add Genetic Complexity
The exceeding rarity of beneficial mutations and the problem this creates for evolutionary time-frames is bad enough. But the next problem evolution faces is insurmountable and it is this: Not a single one of those one-in-a-hundred thousand beneficial mutations can add any significant genetic complexity to an organism! Let me explain.
The majority of mutations fall into two basic classes. They will either jam a genetic signal in the “off” or “on” positions. This is due to the receptor being removed and no longer responding to its signalling molecules. While this may at times be useful, it does not support the assumption that complex biochemical genetics has arisen via evolution. It is clearly a loss of information from the original genetic instruction. The vast majority of these mutations create dysfunction. In extremely rare situations they can benefit an organism. But this is not a mechanism that can create men and women out of a single cell.
For over 100 years, an army of scientists have searched diligently for any mutations that have added beneficial complexity to a genome. Yet, in all of the world’s scientific literature there is not yet a single, clear cut example of a mutation actually adding extra beneficial complexity to an organism. Over and over again I have read in my background study for this essay that their rarity precludes their scientific study. Dr Jerry Bergman’s extensive literature review in 2005 found 453,732 “mutation” hits, but only 186 included the term “beneficial mutation”, about 4 in every 10,000. Of those 186 “beneficial mutations” he found not a single one that unambiguously created new, more complex genetic information. For evolution to exist, it must continually create vast amounts of new genetic information of a higher order, but there is no observable scientific evidence to that effect.
So how can a mutation be beneficial without adding genetic information?
The often quoted examples of beneficial mutations such as adaptive immunity, nucleotide re-combination, antibiotic resistance in bacteria, lactose tolerance in Europeans, resistance to HIV, and sickle-cell anaemia all fail to meet the requirements of an “increasing genetic complexity” mutation. Tyhese examples and all other examples of mutation-enhanced function quoted in the scientific literature involve loss, jamming or mere duplication of genetic information. No novel, more complex genetic information has been added to those organisms. Beneficial mutations are therefore like scratches on a car that accidently make it more aerodynamic; or the same car losing a part so it is lighter and can go faster. The complete lack of examples is strong testimony against the validity of the prime axiom.
This makes perfect sense as informational copying errors such as point mutations, duplications, omissions, deletions, insertions, translocations and inversions can never increase the quality of the original information. This is especially the case as the human genome carries several different messages inside the same single block of DNA, as mentioned above. Our DNA is a multi-dimensional, multi-linguistic language, something we humans will never achieve. Copying errors can never improve an encyclopaedia, a computer code or an instruction manual. DNA is all of these combined, and much, much more.
Evolutionary theory needs a high rate of beneficial mutations and for those mutations to create new information over time. Both of these assumptions are now clearly false. There are virtually no beneficial mutations, those that exist create no significant new information, and there is not enough time. The prime axiom of evolution; that mutations lead to upward genetic progression, is therefore a lie.
But it gets even worse…
3. The Deleterious Mutational Load is Building Rapidly
Exactly how many deleterious mutations are accumulating inside humanity each generation? Now that we have established the truth about the 100,000:1 ratio of deleterious to beneficial mutations and complete lack of genetic progress in the infinitesimal number that are beneficial, it is time to look at the other side of that ratio, that of increasing mutational load. How quickly are the deleterious mutations accumulating in our genomes over time, and is this accumulation a danger to our future existence?
Let’s start by stating that mutation rates affect different species differently. Single-celled asexually reproducing organisms tend to have far lower genetic complexity so obviously experience fewer mutations per individual. E. coli has 4.8 million nucleotides while humans have 3 billion. E. coli is a single cell while we have between 50 and 100 trillion cells. Natural selection is also extremely severe in these “simple” species as every single cell is independently subject to natural selection after every single cell division.
Therefore mutations are a much larger problem in sexually reproducing larger species of trillions of cells and billions of reproductive cells which divide many times before reproduction. The potential for something to go wrong rises exponentially with time. In addition, larger sexually reproducing organisms have a greater accumulation of mutations in the male reproductive cells before reproduction, because of their Y chromosome. Mutations double every 16.5 years in human males as they age, resulting in 76% of all mutations coming down through the paternal line.
Many larger animal species have the additional problem of a relatively low population size with a highly complex genome. Humans have a different problem; a large population but a very low reproductive rate. Finally, the larger the species, the smaller overall effect of each individual mutation on the individual and the less likely it is to be removed via natural selection. In these species, including us, the environment and homeostasis will have a far greater influence on reproduction than natural selection.
The Nobel Prize winning grandfather of modern human genetics, Herman Muller established in 1950 that a mutational load of 0.3 mutations per individual per generation was the limit of human mutational tolerance. The logic was simple. If we have three children per family, we can only afford one of them to carry a large increase in mutational load, and if all carried mutations then at low this level they could easily be selected out. If all our children had mutational loads above this level then they could not be eliminated from the human race. Mutations would begin to accumulate in a linear fashion over time. Instead of evolving upward, the human race would be on a one way trip to eventual extinction.
In 1971 fellow Nobel Prize winner, Manfred Eigen, also calculated that the maximum number of mutations allowable for evolution to progress as 1/n, or one per genome. Any figure above this would eventually result in genetic “error catastrophe”, a term he coined. For many years geneticists, such as James Crow have continued to worry about the effect of increasing numbers of deleterious mutations are having on the human population, particularly with the trend toward older parenting.
So, how many mutations per person per generation are we actually producing? Is it still within the confines needed for evolutionary theory to work suggested by Muller? Advanced studies on the human genome have now shown us the true figure. Sadly, we now know that the single point mutations (SNV’s) alone, without even counting the many other types of mutations, are accumulating on average at between 75 and 175 in our reproductive cells per person, per generation!
This is a profound discovery with huge ramifications for the future of humanity. Because this astounding fact is foundational to the thesis of this essay I have quoted the following admissions from evolutionary geneticists to these mutation rates in humans:
“The average mutation rate was estimated to be approximately 2.5 x 10(-8) mutations per nucleotide site or 175 mutations per diploid genome per generation. The authors find this figure hard to reconcile with evolutionary theory and suggest a mutual cancelling out of mutant nucleotides via epistasis.”
“Recent genome-wide studies of the SNV mutation rate in humans have started to converge. Studies based on whole-genome sequencing and direct estimates of de novo mutations give an average SNV mutation rate of 1.16 × 10−8 mutations per base pair per generation.”
In plain English this estimate is about 30 new SNV’s per person per generation. Table one of their paper gives a mean mutation rate of 96.3 per person per generation. They go on to say that this is a “lower boundary” estimate and that…
“Notably, when considering the total number of mutated base pairs between SNVs and CNVs, CNVs account for the vast majority. CNV’s being copy and deletion mutations covering large number of nucleotides. Put together these two sources of mutations represent hundreds of new mutations per generation.”
“The implication, if these exon rates can be generalized, is of approximately equal to 20 nucleotide mutations per gamete per generation. This estimate of the frequency of point mutations does not include small duplications, rearrangements, or deletions resulting from unequal crossing-over, transcription errors, etc.”
“Every time human DNA is passed from one generation to the next it accumulates 100–200 new mutations, according to a DNA-sequencing analysis of the Y chromosome.”
Many more statements of concern about increasing mutational load by leading human geneticists can be found in Section Six of this essay.
There you have it from some of the world’s leading human geneticists. We are mutating at a rate 100-1,000 times faster than Herman Muller’s upper limit for evolution to be a plausible explanation for the origin of diversified life on this planet. However, these numbers are still not high enough as these quotes only briefly mentioned several of the eight types of mutational damage that is occurring to our DNA. Here are the numbers for some of the others.
Grant Southerland estimates there are as many micro-satellite DNA regional mutations as point mutations. This will add another hundred mutations to the figure of 75-175. Deletion and insertion mutations are relatively few but cause much more damage as they affect more nucleotides at once. They raise the number of mutations per person per generation by another 100. Duplications contribute between 2 and 6 more mutations and are often large scale in nature. Inversions and translocations raise the figure astronomically higher. One recent study put the total figure at around 300 mutations per person per generation. So, the world’s leading human geneticists are telling us that the real mutation rate must therefore be somewhere between 100 and 1,000 mutations per person, per generation. What’s more, most of these mutations are tiny, so are invisible to natural selection. They are therefore accumulating at this rate in every person, every generation in a linear fashion, like spelling mistakes building up in each reprint of an instruction manual, and only one in a 100,000 is beneficial.
But there’s more. Mitochondrial and male Y chromosome mutations are of special concern. Mitochondria mutations add about one mutation per person per generation. They aare particularly significant as mitochondrial replication is a form of asexual reproduction going on within every cell of our body and subject to an inexorable, ever-increasing load of mutations, forever. This fact was first highlighted by Herman Muller and is called Muller’s Ratchet. Commenting on mitochondrial mutations, geneticist Neil Howell suggested in 1996 that “We should increase our attention to the broader question of how (or whether) organisms can tolerate, in the sense of evolution, such a high mutational burden.” The human male Y chromosome also undergoes asexual replication and is therefore subject to Muller’s Ratchet. Rapid mutational build in the Y chromosome is a major cause of falling fertility rates globally. These two asexually reproducing parts of the human body are therefore like rusting cars, ongoing deeply embedded mutational corrosion can only get worse, not better.
In summing up, we now know that the accumulation of all these mutations has resulted in 3.6 million nucleotides of the human genome now becoming corrupt. If genetic entropy is true we would expect to see higher quality genomes the further we go back into history. This is exactly what we find. Recent sequencing of the genomes of a male palaeo Eskimo that lived 4,000 years ago shows he had around 450,000 nucleotide mutations. Such rapid deterioration of the human genome is clear evidence of genetic entropy. It is also clear evidence for a perfect human genome around 4,000BC.
But it gets far worse…
4. The Myth of the Neutral Mutation
With a human mutation rate firmly established by world-renowned researchers at between 100 and 1,000, with two types of asexual duplication taking place within us, and with only one in a 100,000 mutations of benefit to us, the only hope for humanity is that these mutations are not actually affecting our genome. If they are somehow neutral in their effect then humanity would have some chance, however remote, of survival and evolutionary advance.
It is significant to see much of the scientific literature at present engaged in a huge debate as to how much of our DNA is functional and non-functional (junk DNA), and by extension, how many mutations are deleterious or neutral in their effect on genetic fitness. This area seems to be a desperate line in the sand for evolutionists, hoping to rescue their belief that most of our mounting mutational load will not cause significant damage to our genome simply because most of it is non-functional.
At the turn of the 21st Century, before widespread genomic sequencing, it was fashionable to support the notion that, because only 3% of the human genome codes for protein production, the rest must be leftover junk DNA from previous evolutionary episodes. This then meant that 97% of mutations would be absolutely neutral in their genetic fitness effect, allowing evolutionary theorists to breathe a sigh of relief.
However, belief in junk DNA is disappearing rapidly thanks partly to the massive $400 million ENCODE research program which found that about 80% of our genome was functional. Countless other studies around the world are also rapidly advancing our knowledge of the human genome. With every advance in genetics we find ever more functionality in our genome in areas where we thought there was none.
Examples of this ever increasing functional DNA include; Pseudogenes that make regulatory RNA molecules, “selfish genes” that are involved in chromosomal pairing, repetitive and satellite DNA that is heavily involved in genomic function, epigenetic expression, which all comes from non-protein coding regions, and introns that are housed between protein coding sequences and are crucial to accurate gene expression.
As mentioned above, we now know that all DNA is also poly-functional. This means it carries multiple messages in a single sequence. Most DNA will code for RNA on both sides of the double helix. In some cases both strands are transcribed, and in opposite directions. Some DNA sequences code for up to seven different RNA’s from the same length of DNA depending on where transcription is initiated along the code. Other DNA code will sequence for alternate RNA’s depending on the inclusion or exclusion of exons during transcription, or during splicing after transcription. Still other DNA sequences code for both protein binding and protein coding at the same time. The probability of a beneficial mutation is therefore inversely related to the degree to which it was optimised for its original function, and drastically reduces as the number of overlapping codes increases.
On top of all this new-found knowledge of functionality in the DNA structure, gene expression is a field we are only now beginning to understand. Processes such as “imprinting”, “post-translational modification”, “enzyme induction” and “methlaytion” are rapidly expanding our research vocabulary as we learn more about this hugely complex field. The more we learn, the more we realise that even the simplest mutation has to be deleterious to some degree, even if ever so slightly.
It is now obvious to leading researchers that we are no longer dealing with a single DNA code, but many DNA codes. It is not a double helix ladder, but more like a labyrinth. Because of DNA sequencing, the scientific community is falling down Alice’s genetic rabbit hole. So it no longer makes sense to argue that each strand of DNA evolved by mutations operating a few nucleotides at a time. There is absolutely no benefit from any single mutation until a multitude of recombination signals are present, coordinated, functioning and superior to what existed previously. All known forms of DNA mutation championed by evolutionary theory simply disrupt and destroy this poly-constrained coding capacity. It is impossible that mutations could ever create new information in such a hideously complex code operating at near perfection just one level above the atom.
If current trends continue, researchers will soon discover poly-functionality in the entire human genome. Therefore, if there is no junk DNA and all of it is poly-functional, then there is no such thing as a neutral mutation. All mutations will have a deleterious effect. But most of them, being single point mutations will have an ever so slight effect on genetic fitness, but none on reporduction. A smaller number will be minor, visible but not stopping reproduction. A rare few will be major, stopping reproduction altogether. Most will be nearly-neutral, tiny mistakes equivalent to a comma or a single letter on a book-length manuscript, able to slip through the cell’s formidable proof-reading systems. Unless effected by genetic drift, near-neutral and minor mutations will accumulate every single generation, forever. There are no neutral mutations, much to the frustration of evolutionary geneticists who continue to use models assuming most mutations are neutral or beneficial.
But can it get worse…yes!
5. Just How Serious is Human Mutational Load?
Having dealt with the now false idea of neutral mutations, the question now becomes: How large will the cumulative long-term effect of the minor and near-neutral deleterious mutations be, especially in humans?
If 100-1,000 mostly near-neutral reproductive mutations are added to a human genome every generation, they will actually cause little damage to the species initially. Because there are around 3 billion nucleotides in the entire genome, the impact on each generation is linear, not exponential. This slow, inexorable loss of information via tiny mutational accumulation is exactly what we are seeing in the human genome at present, and is of great concern to many of our human geneticists.
Eventually though, after a thousand generations or more, these mutations will begin to cause great damage, just as the repeated photocopying of the copy of a sheet of writing a thousand times will do great damage to the readability of the print. The process is also akin to adding 100-1,000 spelling mistakes to an entire encyclopaedia every annual reprint for a thousand years. For the first five hundred years the text is still readable, but eventually catastrophic breakdown occurs in the message. Indeed, mutational load (genetic entropy) is having a significant effect already in the medical profession where all but a handful of mutations are considered a disaster for the individual. The World Health Organisation estimates that 10,000 diseases are now known to be caused by point mutations in reproductive DNA. The number is increasing annually. The term “neutral mutation” is, by definition, an oxymoron.
As mentioned above, Hermon Muller was the first to recognise the one-way downhill genetic degradation of genomes of all species that reproduce asexually. To avoid the same fate for sexually reproducing species, Muller assumed, hopefully, that the sexual recombination of genes in higher order species would avoid this problem. However, we now know that Muller’s Ratchet applies to all our human mitochondria and male Y chromosomes, as they replicate asexually.
This fact, combined with the number of mutations accumulating inside the reproductive cells of every one of us is now known to be thousands of fold higher than what Muller thought, strongly suggests Muller’s Ratchet also applies to all larger sexually producing species of life on earth. This dire problem has led human geneticist Alexey Kondrashov to title a 1995 research paper: Contamination of the genome by very slightly deleterious mutations: why have we not died 100 times over? The inference is clear; if we are millions of years old as a species we would have died out long ago.
By the way, Muller’s Ratchet does apply to every non-reproductive somatic (normal) cellular division in your body. That’s why we die. Each time your normal somatic cells divide they add between one to three new mutations to the cell. This ever-increasing mutational load is the prime reason why we age and die around 60-90 years. What happens to us individually is what is slowly going to happen to humans, and every other species, in the future.
On that note it is worth mentioning that cloning, taking a somatic cell and using it to produce an exact copy of an individual, is always a genetic disaster. This is because all the previously accumulated mutations in that somatic cell are passed on to the new individual. The clone is born at the age of the donor cell’s giver. The individual will then age and die rapidly as their somatic cells continue this division and mutation process as if they were already middle-aged. It is actually a form of asexual reproduction, therefore Muller’s Ratchet is directly applicable to this procedure. Because the speed of cellular division, and mutational build, is up to 10 fold faster in somatic cells than reproductive cells, cloning is like fast-forwarding genetic entropy 5-10 generations in one!
In conclusion, you can now see the indisputable reality that mutations cannot drive evolutionary advance. Follow the logic: DNA is the rarest and most exquisite of pristine information codes in the universe. The ratio of deleterious to beneficial copying errors in this system is a ratio of a 100,000 to one. There is not enough deep time for those rare beneficial mutations to accumulate. When those exceedingly rare beneficial mutations arise, they do not add any genetic complexity. All the 100-1,000 deleterious mutations building up every generation are mostly near-neutral and invisible to selection. There is no junk DNA to hide near-neutral mutations. Genetic entropy is inevitable. We are not evolving, we are degenerating.
And it gets far worse in Part Three where we discuss the inadequacies of natural selection!