
The evolutionary solution to this massive and relentless accumulation of mutations is to invoke the concept of natural selection as a filter that ensures the elimination of deleterious mutations and the fixing of beneficial mutations. This ensures the survival of the genetically fittest in a species. Natural selection is the second half of the evolutionary prime axiom, and Charles Darwin’s fantasy. However, as just mentioned, natural selection is always and forever a loss of information. It is no friend of evolution. The incorporation of natural selection into the theory of evolution was a desperate solution to an insurmountable Twentieth Century problem. Let me explain.
During the 1930’s, just as Mendelian genetics was being rediscovered, evolutionary theory was embarrassed to find it still held to Darwin’s Lamarckian concepts of change. Darwin did not actually have a theory of inheritance but believed in Lamarckism, the idea that if a giraffe stretched its neck then its offspring would have longer necks. To illustrate, here is a quote from a joint paper Darwin wrote with Alfred Wallace in 1858: “Selection acts only by the accumulation of slight or greater variations, caused by external conditions, or by the mere fact that in generation the child is not absolutely similar to its parent”…” Variations caused by external conditions is Lamarckism through and through.
By the 1930’s evolutionists dominated the halls of western academia so atheistic evolution was going to be kept at all costs. Natural selection, which had long duelled with Lamarckism, was the only mechanism allowed to take its place. Evolutionists now welded genetics and evolution together. Thousands of Mendelian inherited genes, including any mutations, must be subject to natural selection, by decree! This would remove all deleterious mutations while beneficial mutations were able to spread in upward evolutionary progress. They made this assumption based on a tiny understanding of genetics which included the assumed bell curve mentioned earlier.
To convince the general public that inheritance through thousands of tiny genes and natural selection of individuals as a whole unit were compatible concepts, early geneticists, Ronald Fisher, John Haldane and Sewell Wright cleverly decided that natural selection doesn’t work at the level of the individual (phenotype) but at the level of our DNA (genotype). They artificially and arbitrarily transferred the unit of selection from the individual to every individual nucleotide. Modern genetics was built on this false premise that we are simply a swimming “pool” of 3 billion individual nucleotides waiting to be individually selected, and not whole rabbits, geese, humans, trees and whales reproducing!
However, with the rapidly advancing knowledge of genetics from the 1950’s, natural selection immediately faced major problems. It was difficult to reconcile selection at a phenotype level with the existence of a huge number of separate genetic units. The distance between genotype and phenotype was too vast. It was like looking for pixels, within pixels, within pixels, within pixels on a TV, each shrinking in logarithmic order.
To overcome this ever increasing problem, human geneticists have gone on make other assumptions, such as no linkage between nucleotides, unlimited time for selection, unlimited population size and unlimited selection of nucleotides. This process is well documented in the 1971 book The Origins of Theoretical Population Genetics by William Provine. These assumptions keep evolution believable to uncritical thinkers, like university undergraduates. But in the real world they confront a score of fundamental and insurmountable problems: There is little natural selection down at the nucleotide level, little if any individual nucleotide selectivity unless it is so deleterious as to effect reproduction, there is always limited time, and there are always limited population sizes.
All natural selection can do is slow down the mutational loss of information, it can never halt it. In fact it looks much more like a design feature to deliberately slow down the inevitability of genetic entropy. Here are twelve major problems faced by evolutionary reliance in natural selection:
1. The problem of Selection for Phenotype, not Genotype
This is the elephant in the room. As mentioned above, there is a vast gulf between an individual nucleotide and an individual person. Enormous and complex layers of biological organisational structure separate the two, from the level of instructional molecules, to protein structure, to protein function, to cellular life and communication, to tissues, to organs, to individuals. Natural selection never sees individual nucleotides, only individuals, period! Selecting for an individual nucleotide via evolutionary natural selection almost never happens because it would be like a blind person proof reading an entire set of encyclopedias by looking at the covers. Only the extremely significant mutations, like scratches on the book cover or ripped pages, will be felt by the blind process of natural selection. Selection via genotype instead of phenotype is the great deception our culture is forced to believe in or evolutionary theory unravels.
This one fact of phenotype selection, instead of genotype selection, extinguishes nearly all attempts at mutational selection by evolutionary processes.
2. The Problem of Near-Neutral Mutations
Remember the exponential deleterious mutational curve illustrated previously? Nature dictates that the closer the point on the bell curve to the middle, the more mutations there are. Mutations are always exponentially skewed to this near-neutral zone. The smaller the mistake the more likely it is to slip through. Very few mutations lay further out, visible to selection processes, the unshaded section of the curve. This means that all near-neutral mutations (those that produce no felt effect on the individual) and minor mutations (those that are felt but do not stop reproduction) are totally invisible to natural selection and therefore impossible to remove from the population (the shaded section of the illustration below). Natural selection is utterly and completely blind to what exists inside an organism’s DNA. It is like feeling for a pea under a thousand kilometre high mattress.
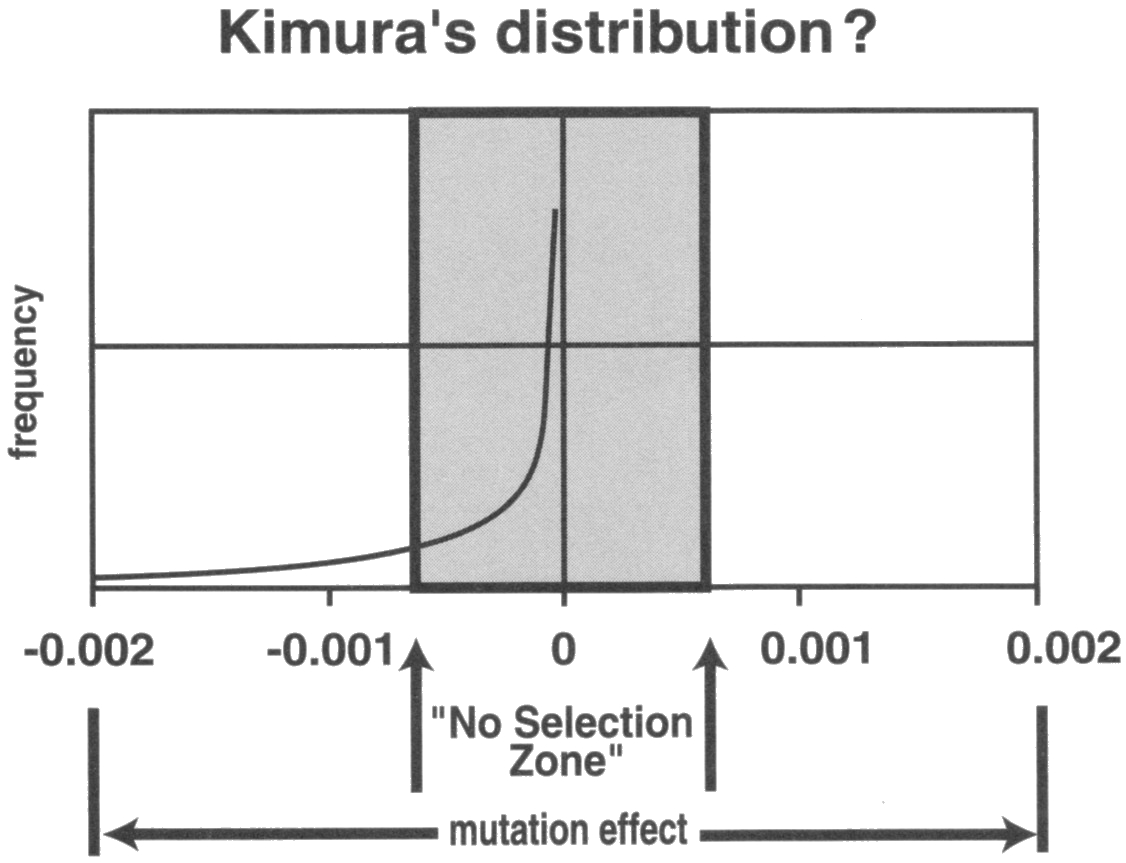
Removal of mutations at reproduction can therefore only occur if they are significant enough to overcome the powerful reproductive urge in all individuals in all species. Because virtually all mutations are near-neutral or minor, and therefore invisible to natural selection, mutational load is destined to build in all species, all the time. This increase is accelerating rapidly in humans since the advent of modern medicine has allowed millions of us to reproduce who would have died before reproducing in previous centuries.
Each one of us is a therefore a slowly ticking genetic time bomb. Genetics is not onward and upward. Near-neutral mutations will only be removed when, after hundreds or thousands of generations, their combined effect on genetic fitness finally impairs reproduction on a mass scale. We call this event extinction.
3. The Problem of Recessive Mutations
To make matters worse, most deleterious mutations are genetically recessive. They do not show up every generation but only when an individual breeds with another individual with the same recessive mutation. They are usually passed on silently. Given that we are generating somewhere between 100 and 1,000 new mutations each time we breed, the human race is collectively accumulating between 600 billion and 6 trillion new mutations every generation. As time progresses the chances of every person mating with another person carrying the same recessive mutation inescapably increases. In acknowledgement of the enormity of this problem evolutionary geneticist James Crow believes the fitness level of the human genome is now dropping at between 1-2% per generation. More recently geneticist Michael Lynch thinks it may be actually dropping at around 5% every generation! Survival of the fittest thus becomes a relative term because in the next generation every individual is less genetically fit than the last. All of us are on a genetic version of the Titanic.
4. The Problem of Selection Cost
The whole rationale of natural selection is…selection! Selection always equals elimination. Nature, all by itself, is meant to eliminate genetically unfit individuals and favour those with superior genes. But there is a cost to any and all selection. Selection means removing less fit individuals from the breeding process, either through death or disability. Therefore nature must always produce a surplus of individuals to account for those that do not reproduce. If it doesn’t, mutational load will increase inexorably and the species is on a one way path to extinction. In addition, most individuals, and even some genes, will have a combination of desirable and undesirable traits. Elimination against the undesirable trait costs a species the opportunity to pass on the desirable trait. Selection of the desirable trait also passes on any undesirable trait. Catch 22.
In the case of humans, our global fertility rate is now 2.36 children per couple. This means at this point in history we have almost zero excess children for selection processes. This problem was first alluded to by John Haldane in 1957 and has been subject to rigorous debate ever since. Many modern geneticists fret over the low reproductive rate of humans and wonder how evolution could proceed in these circumstances. Selected quotes from them are provided in Section Six below.
5. The Problem of Noise
Any type of interference between significant mutational signals and their reproductive visibility is called “noise” as it inhibits the genetic signal. Most noise is environmental. The higher the level of environmental noise, the weaker the natural selection process becomes. Geneticists estimate that 50% of the difference between individual reproductive success rates for all species are due to purely to environmental noise factors; nurture rather than nature. For example, reproductive success for humans is heavily dependent on environmental factors such as social trends, contraception, lifestyle, availability, diet, war, accidents, marriage rates, and disease. In addition to environmental factors, genetic/environmental susceptibility is responsible for another 25% of reproductive success. For example, we can have the gene for breast cancer, but if we live in a clean environment with a great diet it may never develop, and vice versa. For the remaining 25% of the reproductive success spectrum you have the additional problems of mutational recessiveness, epistasis, homeostasis, cyclical selection and epigenetics. More on those terms later…
Many evolutionary websites will assign amazing power to natural selection as if there is a one-to-one correlation between mutations and reproduction. They are playing games with the truth. Biological noise leaves little to no space for mutational advancement. John Sanford’s research shows that noise takes the shaded non-selection zone, illustrated in the exponential mutational curve above, and extends it to the point where even most major deleterious mutations are also subject to non-selection. This is why Kimura (The Neutral Theory of Genetic Evolution, 1983, page 31) estimated that only 1% of genetic fitness is attributable to genetically heritable factors. If it is this low for a gene of between 80 and 80,000 nucleotides, imagine how low it is for a single nucleotide mutation, good or bad!
6. The Problem of Genetic Drift
In a species with a small population, genetic drift also plays a very large role in reproductive success. Random sampling errors in the sexual recombination of chromosomes logically have a greater effect in small populations, just as they do when tossing just a few coins. The smaller the sample size, the less natural selection will work because heritable traits will pop up much more inconsistently. Population geneticists will therefore often assume unlimited populations to get around this problem of genetic drift so their modelling will work. This does not reflect reality. Because of genetic drift, most of the world’s endangered species will be extinct in the not too-distant future when genetic meltdown sets in.
7. The Problem of Large Linkage Blocks
Genetic duplication always occurs in blocks, not individual nucleotides. In humans, nucleotides that are close together are passed on together in linkage blocks averaging 30,000 nucleotides in size. There are about 100,000 such blocks. Our 23,000 genes themselves are another even larger linkage block of duplication, while chromosomes are the largest linkage block. It is only at the level of linkage blocks that genetic traits are passed on, visible to selection and therefore heritable. All individual nucleotide mutations, good or bad, are being passed on in blocks. Their little voice cannot be heard over the top of a crowd of 30,000 others unless it is highly deleterious. Therefore all near-neutral and minor deleterious mutations within that block will be passed on. They can’t be selected out. This is yet another reason why genetic load will forever increase in a Muller’s Ratchet scenario, the opposite of evolutionary theory.
8. The Problem of Invisible Beneficial Mutations
If all near-neutral and minor deleterious mutations are being passed on in linkage blocks and accumulating, then those extremely rare beneficial mutations are also locked inside linkage blocks and cannot be positively selected for individually. No positive natural selection can occur at the nucleotide level! This means zero evolutionary advancement unless a mutation is both monstrously positive and adding to genetic complexity. To observe the first is like accidentally finding a pin floating in interstellar space as it will only occur once in a billion or trillion mutations. The second as we know is impossible as all mutations impede some form of previously higher level information flow. Evolutionists now boldly claim that large scale duplications of entire linkage blocks will solve this problem. This claim is dealt with in detail in Section Seven below.
9. The Problem of Homeostasis
Homeostasis also disrupts natural selection. Homeostasis is our internal regulation system in response to our environment. Sensors in all an organism’s cells send messages to other cells calling for internal adaptation to incoming external stimuli. Our internal environment counteracts external stimuli, stunting natural selection pressures. Examples include growing fur in winter, reducing blood flow in external limbs in cold climates, and sweating in summer heat. This helps us survive when we otherwise may not. Our genetic disposition can be masked by this ability to cope with our environment.
10. The problem of Epistasis
No nucleotide has any meaning or purpose outside its relationship with vast numbers of nucleotides around it, beginning with its three letter codon right up through to chromosomes. They all depend on each other for meaning. They create a living language, just as the letters on this page, and in this entire essay are meaningless unless read in context. We call this epistasis. Because there are 3 billion nucleotides, each one carries an almost infinitely small amount of information. They are almost useless and carry no meaning outside the bigger message. You need a major change to this bigger message before any selection can take place. Any beneficial mutations will be drowned out and invisible to selection unless there were a group of them all occurring at the same time in the same place. Since the ratio of deleterious to beneficial mutations is 100,000:1, the chances of a group of beneficial mutations occurring in one place at one time approaches statistical zero.
11. The Problem of Multiple Selection
Assume for a moment that you did manage to select for any given nucleotide or entire linkage block and it was visible to selection. To select for that one visible trait, you then reduce the ability to select for any other nucleotide or linkage block. This is the dilemma of artificial breeding in plants and livestock. It is give and take. Selecting for more than a few traits becomes a useless exercise. Selection for one discounts another. Selecting for every mutation in each human generation is clearly impossible. Many human geneticists, to make their models workable, conveniently argue that you actually can select for any number of mutations per breeding cycle. Ignoring this reality helps validate their models. However, any plant or animal breeder knows better.
12. The Problem of Epigenetics
Epigenetic deals with non-permanent changes to DNA. These changes are a response to the environment, but they entail switching existing genes off and then on again several generations later. This inbuilt survival mechanism does not involve the creation of new genes, a fact lost on the many websites promoting it as a powerful evolutionary force. In fact epigenetics makes it even more difficult for natural selection to work because non-permanent changes act like a moving target for natural selection. Epigenetics adds much more biological noise to environmental noise, making it that much harder for natural selection to operate. The mutational non-selection zone for natural selection just keeps getting larger and larger.
Conclusion
In conclusion to this section on natural selection, the harsh reality is that there is no evolutionary mechanism capable of genetic advancement, only an ever-increasing genetic load. Natural selection is powerless to eliminate this ever increasing genetic burden. It is crippled by; phenotype selection, the non-selection of near-neutral mutations, the recessiveness of most mutations, the existence of linkage blocks, the invisibility of beneficial mutations, selection costs, multiple selection impossibility, epistasis, epigenetics, homeostasis and genetic drift. All evidence points to a non-selection box that includes all but the few very rare catastrophic mutations. We are all slowly dying out. Natural selection simply doesn’t work in the real world.
In the real world natural selection is a design feature that stabilises a population that would otherwise die out rapidly. It cannot halt an ultimate genetic meltdown but it can slow it down considerably, enabling life to continue on earth for hundreds or thousands of generations longer than otherwise possible.
Part Four of this essay looks into the probability of unguided mutational processes creating a single complete human gene over 6 million years.